Our laboratory is interested in defining the structure
and properties of the unique class of membrane channels called
gap junctions that allow the direct passage of ions, small
metabolites and secondary messengers between cells. The proteins
that comprise these channels, a family called connexins in
the vertebrates , are diverse in nature, with multiple members
of the family being expressed in most cells and tissues. It
has become increasingly evident that this diversity in connexin
composition imparts differential regulatory and permeability
properties to these intercellular channels. Understanding
the structural basis underlying the different properties of
connexins will be an essential step in fully appreciating
the specialized role that these structures play in different
tissues. Evidence that structural diversity has physiological
consequences is provided by the linkage of five very distinct
human diseases to defects in different connexin genes. Specifically,
deafness is linked to Cx26 and Cx31 mutations, a form of skin
keratinopathy is linked to distinct defects in Cx31, peripheral
neuronal degeneration in Charcot Marie Tooth's disease is
linked to a plethora of Cx32 defects and catarracts are linked
to Cx50 defects . Similarly, knock-outs of different connexins
in mice have produced highly variable problems ranging from
embryonic death (Cx26), to increased susceptibility to tumors
(Cx32 ) or cardiac arrhythmias (Cx40), female sterility (Cx37
) and eye catarracts
(Cx46 and 50).
Our own work has contributed significantly
to defining patterns of selective interactions between connexins
that appear to be important in establishing communication
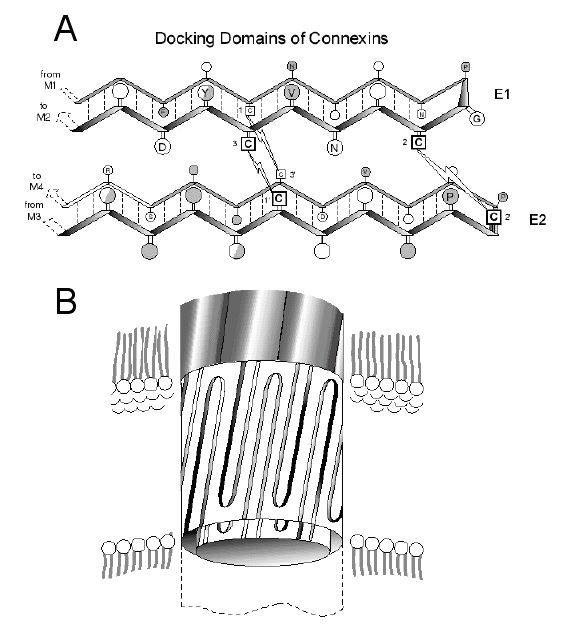
boundaries in vivo. Site-directed mutagenesis, combined with
biochemical and functional analyses of connexins expressed
in oocyte pairs or cell-free systems, has allowed us to define
the structural basis for this "docking" interaction between
connexins of apposed cells
[Foote etal. J. Cell Biol. 140: 1187 (1998)]. Similar
strategies have also been used in probing channel gating mechanisms.
Identification of functional domains that are involved in
the gating of the channels in response to voltage [Suchyna etal. Nature 365: 847 (1993)] , and phosphorylation
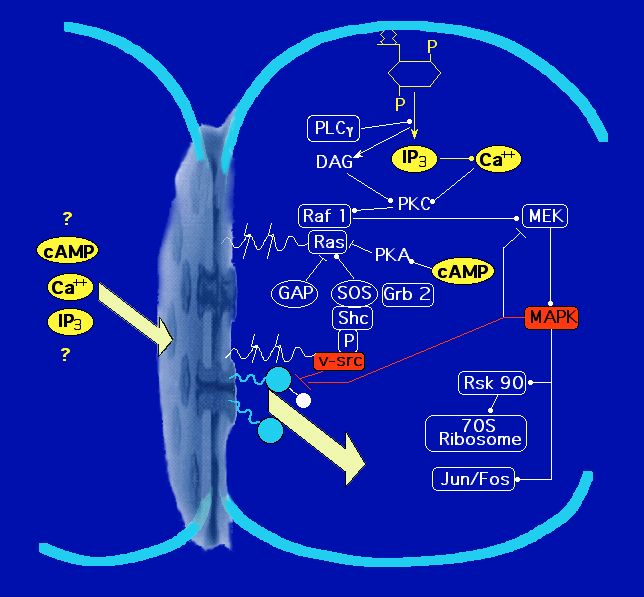
by MAPkinase
[Zhou etal. J. Cell Biol., 144: 1045 (1999)] has indicated
that these processes occur through quite distinct molecular mechanisms. We have also been investigating
the different permeability properties of gap junction
channels composed of different connexins
[Cao etal. J. Cell Science 111: 31 (1998)] . Recently,
this has been extended to the identification of natural metabolites
that pass preferentially through different channels. In order
to identify the determinants for channel selectivity, we have
also been employing the SCAM technique of cysteine scanning
mutagenesis to identify the domains of the protein that contribute to
the channel lining.
The long-term aim of these
studies is to better understand the biological role played
by gap junctions in different systems. A particular focus
is the mechanism by which gap junctions act as
tumor suppessors.
As part of these studies, we are comparing the permeability of
connexins that have proven to be effective growth suppressors
, to those that are not. We have also been investigating the mechanism
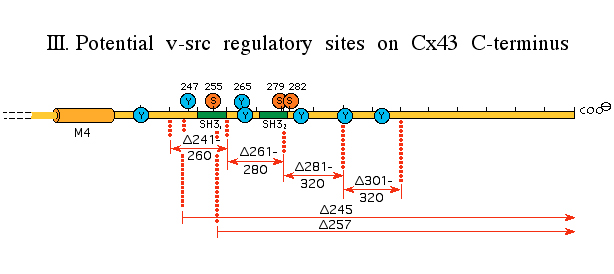
through which some oncogenes
(e.g. v-src) can inhibit coupling
. This work has shown the mechanism to be like the "ball and chain" gating
of K+ channels, in this case instigated by a phosphorylation
event, apparently involving MAPkinase.
[Zhou etal. J. Cell Biol., 144: 1045 (1999)]
This provides us with tools
to selectively prevent the uncoupling of cells by v-src, allowing
the role of gap junctions in inhibiting the transforming effects
of this oncogene to be assessed.